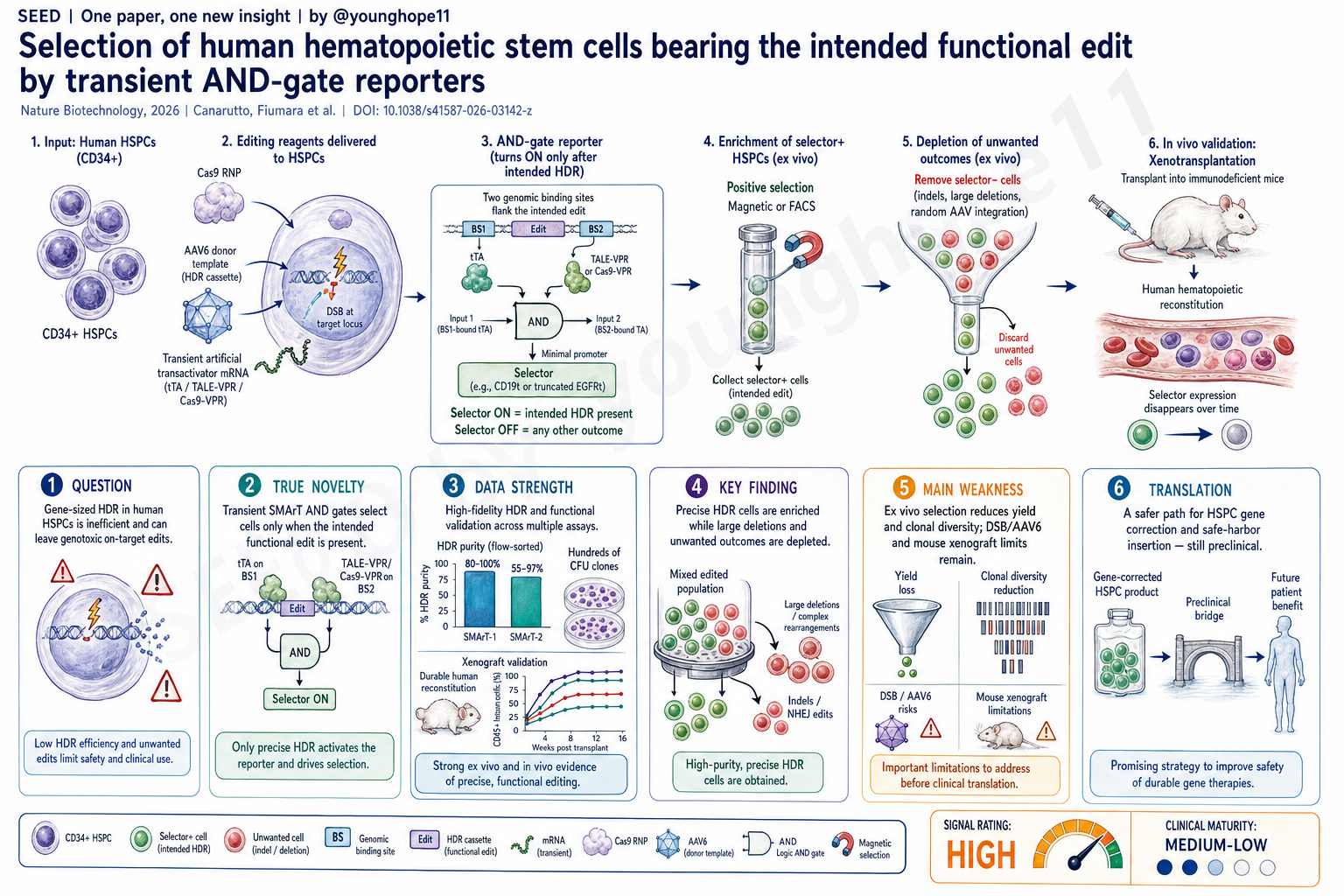
SEED | 给 HSPC 精准 HDR 加一道短暂门控 SEED | A transient gate for precise HSPC HDR AI-assisted · reviewed
San Raffaele Telethon Institute for Gene Therapy 的 Daniele Canarutto、Martina Fiumara 与 Samuele Ferrari、Luigi Naldini 团队近期报道,利用 transient artificial transactivator 构建 SMArT AND-gate reporters,可以在人 HSPCs 中富集携带预期功能性 HDR 编辑的细胞,同时优先剔除潜在 genotoxic on-target edits,为 gene-sized HSPC editing 提供了一条更可控的选择与制造路径。
研究问题是什么?
这篇论文问的是 HSPC gene editing 里一个非常实际的瓶颈:如果我们想把 gene-sized therapeutic cassette 精准整合到目标位点,能不能只保留真正带有预期功能性 HDR 编辑的细胞,而不是把精确编辑、indel、大缺失、模板错误插入和其他 on-target rearrangement 混在同一个治疗产品里?
HDR editing 的吸引力很清楚。它可以修复 disease-causing locus,也可以把表达 cassette 放进 safe harbor,让细胞获得新的治疗功能。但在人 HSPCs 里,HDR 受细胞周期限制,效率不稳定;同时,CRISPR-induced DSB 会产生 NHEJ/MMEJ、large deletion、chromosomal translocation、chromothripsis 或 HDR template trapping 等异常结果。最终产品因此可能是一个遗传上高度异质的细胞群,其中一部分没有疗效,一部分还可能带有位点相关的 genotoxic risk。
所以,真正的问题不是“能不能编辑 HSPC”,而是“能不能在不长期表达 selector、不破坏生理调控、不明显损害 engraftment 的前提下,把 intended functional edit 从复杂编辑产物中选出来”。
真正的新意是什么?
真正的新意是把 selection 设计成一个短暂的 genomic AND gate:selector 不是持续表达的标记物,而是只有当 HDR cassette 按预期整合到目标位点、且 transient artificial transactivator 能在正确上下文中激活它时,才会短暂打开。
作者把这个思路命名为 SMArT,selection by means of artificial transactivators,并设计了三种版本。SMArT-1 把 tTA-responsive selector 放进 AAV6 HDR donor,在非整合模板逐渐稀释后用 tTA mRNA 短暂打开 selector,适合快速验证和较强可移植性。SMArT-2 把 selector 与 corrected cDNA 功能耦合,由目标基因内源 promoter 和人工转录激活器共同驱动,更适合像 IL2RG 这类需要恢复生理表达调控、同时希望剔除 X-linked male cells 中异常 on-target 事件的场景。SMArT-3 则把 selector 放在 HDR template 中的 minimal promoter 下,由结合在 homology arm 外侧的 ArT 激活,避免 selector 长期存在,也不要求目标基因本身适合 transactivation。
这比传统的“带一个 constitutive selector 再排序”更精细。它选择的不是“细胞里有 vector”或“细胞表达了一个永久 marker”,而是“基因组在预期位置形成了可被短暂逻辑门识别的功能性结构”。后续用 Cas9-VPR 和 truncated gRNA 把 SMArT-3 简化成一个 polyfunctional editor,也让这套方法更接近可扩展平台。
数据强在哪里?
强在它不是只做了一个 reporter system,而是跨多个层级验证了“能选中正确编辑、能清除异常产物、还能保留 HSPC 功能”这三个关键点。
第一层是平台广度。作者在 K-562 reporter cells、cord blood HSPCs、mobilized peripheral blood HSPCs 和 T cells 中测试;目标位点包括 IL2RG、CD40LG、AAVS1 和 RAG1 相关设计;selector 也从 GFP 扩展到 CXCR4、ΔLNGFR 和 tEGFR 等更接近临床制造的选择标记。整体上,论文报告 HDR-edited HSPCs 可通过短暂 selector expression 富集到 80-100% purity。
第二层是 SMArT-2 对 IL2RG 的 stringency。T3-VPR transactivation 让 selector expression 平均覆盖约 75% 的 HDR-edited HSPCs;在单克隆 colony 层面,bulk edited cells 从约 55% HDR enrichment 提高到 ΔLNGFR+ fraction 中约 97%。更重要的是,IL2RG promoter 大缺失在 ΔLNGFR+ colonies 中没有检测到,而在 bulk 和 ΔLNGFR- colonies 中仍可见。作者还用 native DNA long-read sequencing 进一步确认 selected fraction 更富集 predicted functional rearrangements。
第三层是功能验证。SMArT-2-selected HSPCs 能在 artificial thymic organoids 中分化到 CD4 或 CD8 single-positive T cell stage,并保留多样 TCR Vβ repertoire。异种移植到 NBSGW mice 后,positive selection group 仍能形成 robust human graft,且在 bone marrow、spleen 和 peripheral blood 中显示约 2.2-3 倍更高的 HDR editing。
第四层是 SMArT-3。AAVS1 位点的 mPB HSPC 实验显示,T7-VPR 可在 bulk treated HSPCs 中诱导中位数约 25% 到 35% 的 GFP expression,在最 primitive CD90+ compartment 中也能达到约 15% 到 25%。分选后的 GFP+ colonies 中,SK 和 mCMV 条件下分别有 100% 和 97% 至少带有一个 HDR-edited allele。异种移植时,SMArT-3-selected cells 产生的 human graft 可达到约 1-1.5 个 HDR-edited AAVS1 alleles per diploid genome,接近 fully edited graft,而且 selector expression 随时间消失。优化 GMP-compatible culture、electroporation 和 sorting 后,selected graft 的 engraftment 与 bulk cells 相当,同时仍保持平均约一个 edited allele per diploid genome。
最大弱点是什么?
最大弱点是 selection 本身会带来制造和生物学代价。最直接的风险是 yield 和 clonal diversity。早期 SMArT-3 xenograft 中,GFP+ selected group 的 graft size 比 bulk 或 GFP- group 低 5-10 倍;虽然优化流程后 engraftment 明显改善,但这也说明 HSPC editing、AAV6 exposure、electroporation、sorting 和 selection window 共同构成了真实制造压力。
第二个限制是安全检测仍需要更深。作者自己指出,targeted ddPCR 是 HDR 的有用 surrogate,但它对更复杂的 genomic rearrangements、imprecise targeted integration、vector concatemer trapping 等结果并不敏感。论文加入了 native long-read sequencing,这是一个强点;但若要走向临床,还需要更系统的 off-target/on-target genotoxicity profiling、clonal tracking、barcode-based diversity measurement 和更长期的 tumorigenicity 风险评估。
第三个限制是平台适用性不是无限的。SMArT-2 在 IL2RG 这样的 X-linked male setting 中特别有力,因为 selection 可以接近完全剔除某些大缺失;但在 autosomal loci 或女性 X chromosome 上,不能预期同样彻底地清除所有异常等位基因。SMArT-3 更通用,但可能牺牲 selection yield,而且强 promoter 或某些基因组上下文可能导致 unwanted basal activation。
第四个限制是 cargo 和流程复杂度。AAV6 cargo size 仍是硬约束;ΔLNGFR selector 本身约 843 bp,会挤占 therapeutic cassette、homology arms 和调控元件空间。整个流程也不仅是“多加一个 marker”,而是 RNP/AAV6/ArT mRNA/selector/sorting/GMP manufacturing 的系统工程,需要证明复杂性换来的安全收益足以覆盖成本和制造风险。
是否有临床转化意义?
有,而且主要是平台层面的转化意义。它面向的是 ex vivo HSPC gene therapy 中最难解决的一类问题:如何把 gene-sized edit 做得更纯、更可控、更少 genotoxic burden,同时保留长期造血重建能力。
对 inborn errors of immunity,IL2RG、CD40LG、RAG1 这类场景尤其相关。与 lentiviral gene addition 相比,精准 gene correction 的潜在优势是更接近生理表达调控;而 SMArT 的价值在于把正确修复的细胞富集出来,并减少带有潜在危险 on-target edits 的细胞进入最终产品。对 safe harbor insertion 或 future cancer gene therapy,SMArT-3 和 Cas9-VPR polyfunctional editor 也提供了更可移植的路线。
但它离临床应用还不是一步之遥。当前证据主要来自体外人 HSPC、T cell、colony assay、ATO 和 immunodeficient mouse xenograft。下一步需要在 patient-derived HSPCs、larger-scale GMP runs、clinically realistic cell dose、long-term engraftment、barcode-based clonal output 和 regulatory-grade genotoxicity assay 中证明同样成立。
Yang 的信号评级:High
理由:我会把这篇评为科研信号 High,因为它不是单纯提高 HDR 百分比,而是提出了一套把“正确编辑”转化为“可选择细胞状态”的平台逻辑。SMArT 的核心贡献,是把 functional genomic outcome、transient reporter expression、cell sorting 和 genotoxic burden reduction 连接成同一个制造框架。
它的数据也支撑这个 High:多种 SMArT 架构、多种 selector、多种位点、人 HSPCs、single-colony genotyping、long-read sequencing、T cell differentiation 和 mouse xenotransplantation 之间形成了比较完整的证据链。弱点并不小,尤其是 yield、clonal diversity 和复杂制造,但这些弱点没有推翻中心结论。
但这个 High 是“科研信号 High”,不是“临床就绪 High”。临床成熟度我会单独评为 Medium-Low:它已经明显比概念性 reporter 更接近可制造平台,也使用了 clinically compliant selectors 和流程优化;但还没有患者级疗效、安全性和规模化制造数据,真正进入临床仍需要严肃的工艺与监管验证。
At the San Raffaele Telethon Institute for Gene Therapy, Daniele Canarutto, Martina Fiumara and the team of Samuele Ferrari and Luigi Naldini recently reported SMArT, a transient artificial-transactivator AND-gate reporter strategy that enriches human HSPCs carrying the intended functional HDR edit while preferentially depleting potentially genotoxic on-target outcomes, providing a more controllable selection and manufacturing path for gene-sized HSPC editing.
What is the research question?
This paper asks a very practical question in HSPC gene editing: if we want to integrate a gene-sized therapeutic cassette precisely into a target locus, can we keep only the cells carrying the intended functional HDR edit instead of mixing precise edits, indels, large deletions, erroneous template insertions and other on-target rearrangements in the same therapeutic product?
The appeal of HDR editing is clear. It can repair a disease-causing locus, or place an expression cassette into a safe harbor so that a cell gains a new therapeutic function. But in human HSPCs, HDR is constrained by cell cycle state and remains inefficient. At the same time, CRISPR-induced DSBs can produce NHEJ/MMEJ, large deletions, chromosomal translocations, chromothripsis or HDR template trapping. The final product can therefore become a genetically heterogeneous cell population in which some cells may be ineffective and others may carry locus-specific genotoxic risk.
The real question is therefore not just whether HSPCs can be edited. It is whether the intended functional edit can be selected out of a complex editing mixture without permanent selector expression, without disrupting physiological regulation and without substantially damaging engraftment.
What is truly new?
The real novelty is the design of selection as a transient genomic AND gate. The selector is not a permanently expressed marker. It turns on only when the HDR cassette has integrated as intended at the target site and a transient artificial transactivator can activate it in the correct genomic context.
The authors call this strategy SMArT, selection by means of artificial transactivators, and build three versions. SMArT-1 places a tTA-responsive selector into the AAV6 HDR donor; after non-integrated template DNA is diluted, tTA mRNA transiently activates the selector, making this version relatively portable and useful for rapid validation. SMArT-2 functionally couples the selector to the corrected cDNA and uses the endogenous target-gene promoter plus an artificial transactivator, which is better suited for settings such as IL2RG where physiological regulation should be restored and unwanted on-target events in X-linked male cells should be depleted. SMArT-3 places the selector under a minimal promoter embedded in the HDR template and activates it using an ArT that binds outside the homology arm, avoiding permanent selector expression and removing the need for the target gene itself to be easily transactivated.
This is more precise than adding a constitutive selector and sorting. It does not simply select cells that contain vector DNA or express a permanent marker. It selects cells whose genome has formed a functional structure that can be recognized by a transient logic gate. The later Cas9-VPR and truncated-gRNA implementation also makes SMArT-3 feel closer to a scalable platform.
Where is the data strongest?
The strength is that the paper does not stop at one reporter system. It tests three linked claims across multiple levels: correct edits can be selected, unwanted outcomes can be depleted and HSPC function can be preserved.
The first layer is platform breadth. The authors test K-562 reporter cells, cord blood HSPCs, mobilized peripheral blood HSPCs and T cells. The target loci include IL2RG, CD40LG, AAVS1 and RAG1-related designs. The selectors also extend from GFP to more clinically relevant options such as CXCR4, Delta LNGFR and tEGFR. Overall, the paper reports that HDR-edited HSPCs can be enriched to 80-100% purity through transient selector expression.
The second layer is the stringency of SMArT-2 at IL2RG. T3-VPR transactivation allowed selector expression to cover an average of about 75% of HDR-edited HSPCs. At the single-colony level, bulk edited cells went from about 55% HDR enrichment to about 97% in the Delta LNGFR-positive fraction. More importantly, large deletions encompassing the IL2RG promoter were not detected in Delta LNGFR-positive colonies, while they remained detectable in bulk and Delta LNGFR-negative colonies. Native DNA long-read sequencing further confirmed enrichment for predicted functional rearrangements in the selected fraction.
The third layer is functional validation. SMArT-2-selected HSPCs differentiated in artificial thymic organoids into CD4 or CD8 single-positive T cell stages and retained a diverse TCR V beta repertoire. After xenotransplantation into NBSGW mice, the positive-selection group still generated robust human grafts and showed about 2.2- to 3-fold higher HDR editing in bone marrow, spleen and peripheral blood.
The fourth layer is SMArT-3. In mobilized peripheral blood HSPCs at the AAVS1 locus, T7-VPR induced median GFP expression of about 25% to 35% in bulk treated HSPCs and about 15% to 25% in the primitive CD90-positive compartment. After sorting, 100% and 97% of GFP-positive colonies in the SK and mCMV conditions carried at least one HDR-edited allele. In xenografts, SMArT-3-selected cells produced human grafts with about 1 to 1.5 HDR-edited AAVS1 alleles per diploid genome, approaching a fully edited graft, while selector expression disappeared over time. After optimization of GMP-compatible culture, electroporation and sorting, engraftment of selected grafts became comparable to bulk cells while maintaining an average of about one edited allele per diploid genome.
What is the biggest weakness?
The biggest weakness is that selection itself creates manufacturing and biological costs. The most direct risk is yield and clonal diversity. In the early SMArT-3 xenograft experiment, the GFP-positive selected group had 5- to 10-fold lower graft size than the bulk or GFP-negative groups. Optimization improved engraftment substantially, but this also shows that HSPC editing, AAV6 exposure, electroporation, sorting and the selection window together impose real manufacturing stress.
A second limitation is that safety detection still needs to go deeper. The authors note that targeted ddPCR is a useful surrogate for HDR, but it is blind to more complex genomic rearrangements, imprecise targeted integration and vector concatemer trapping. The addition of native long-read sequencing is a strength, but clinical translation would still require more systematic off-target and on-target genotoxicity profiling, clonal tracking, barcode-based diversity measurement and longer-term tumorigenicity risk assessment.
A third limitation is that platform applicability is not unlimited. SMArT-2 is especially powerful in an X-linked male setting such as IL2RG, where selection can nearly eliminate certain large deletions. But at autosomal loci or on the female X chromosome, one should not expect the same exhaustive removal of all abnormal alleles. SMArT-3 is more general, but may trade off against selection yield, and strong promoters or certain genomic contexts could create unwanted basal activation.
A fourth limitation is cargo and process complexity. AAV6 cargo size remains a hard constraint. The Delta LNGFR selector itself is about 843 bp, reducing the remaining space for the therapeutic cassette, homology arms and regulatory elements. The workflow is also not just the addition of a marker. It is a system involving RNP, AAV6, ArT mRNA, selector design, sorting and GMP manufacturing, and the safety benefit will have to justify that added cost and process risk.
Is there translational or clinical relevance?
Yes, mainly at the platform level. The work addresses one of the hardest problems in ex vivo HSPC gene therapy: how to make gene-sized edits purer, more controllable and less burdened by genotoxic outcomes while preserving long-term hematopoietic reconstitution.
For inborn errors of immunity, settings such as IL2RG, CD40LG and RAG1 are especially relevant. Compared with lentiviral gene addition, precise gene correction can potentially restore more physiological regulation. The value of SMArT is that it enriches correctly repaired cells and reduces the entry of cells with potentially dangerous on-target edits into the final product. For safe-harbor insertion or future cancer gene therapy, SMArT-3 and the Cas9-VPR polyfunctional editor also offer a more portable path.
But this is not one step away from clinical use. The current evidence is mainly from ex vivo human HSPCs, T cells, colony assays, artificial thymic organoids and immunodeficient mouse xenografts. The next step is to prove the same logic in patient-derived HSPCs, larger-scale GMP runs, clinically realistic cell doses, long-term engraftment, barcode-based clonal output and regulatory-grade genotoxicity assays.
Yang’s signal rating: High
Reason: I would rate this paper’s scientific signal as High because it does not merely increase the percentage of HDR. It proposes a platform logic that turns the correct edit into a selectable cell state. The core contribution of SMArT is to connect functional genomic outcome, transient reporter expression, cell sorting and genotoxic-burden reduction into one manufacturing framework.
The data also support that High rating. Multiple SMArT architectures, multiple selectors, multiple loci, human HSPCs, single-colony genotyping, long-read sequencing, T cell differentiation and mouse xenotransplantation together form a fairly complete evidence chain. The weaknesses are real, especially yield, clonal diversity and manufacturing complexity, but they do not overturn the central claim.
But this High means scientific signal High, not clinical readiness High. I would separately rate clinical maturity as Medium-Low: the work is clearly closer to a manufacturable platform than a conceptual reporter and it uses clinically compliant selectors and process optimization, but it still lacks patient-level efficacy, safety and scaled manufacturing data. Clinical translation will require serious process and regulatory validation.