SEED | Prion 里藏着抗菌肽 SEED | Prions hide antimicrobial peptides AI-assisted · reviewed
University of Pennsylvania Machine Biology Group 的 Marcelo D. T. Torres、Fangping Wan 与 Cesar de la Fuente-Nunez 团队近期报道,通过 APEX 1.1 深度学习框架系统扫描 prion 和 prion-like proteins,发现其中隐藏着一类具有抗菌活性的 encrypted peptides,并将其命名为 prionins;这为从非典型蛋白类别中挖掘新抗菌肽提供了新的来源空间和验证路径。
研究问题是什么?
这篇文章问的是一个反直觉问题:prion 和 prion-like proteins 是否只与蛋白错误折叠、amyloid formation 和神经退行性病变相关,还是也可能在一级序列中藏有可被释放、剪切或人工挖掘出来的 antimicrobial peptides?
这个问题不是凭空来的。已有研究显示 amyloid-beta、cellular prion protein 等 amyloid-associated proteins 可以表现出 antimicrobial 或 host-protective activity。因此,作者把 prion biology 和 encrypted peptide antibiotic discovery 连接起来:如果许多非免疫蛋白都能藏有抗菌肽,那么聚集倾向强、结构特殊的 prion-related proteome 也可能是一座未被系统挖过的 peptide mine。
更具体地说,作者要回答三件事。第一,prion-related proteins 中是否存在大量可被 AI 预测为抗菌的短肽片段?第二,这些片段是否真的能在体外抑制临床相关细菌,并呈现可解释的膜扰动机制?第三,少数 lead peptide 是否能在动物感染模型中产生真实抗感染效果?
真正的新意是什么?
真正的新意是把 prion 从“病理蛋白类别”重新定义成 antibiotic discovery 的 sequence source space。
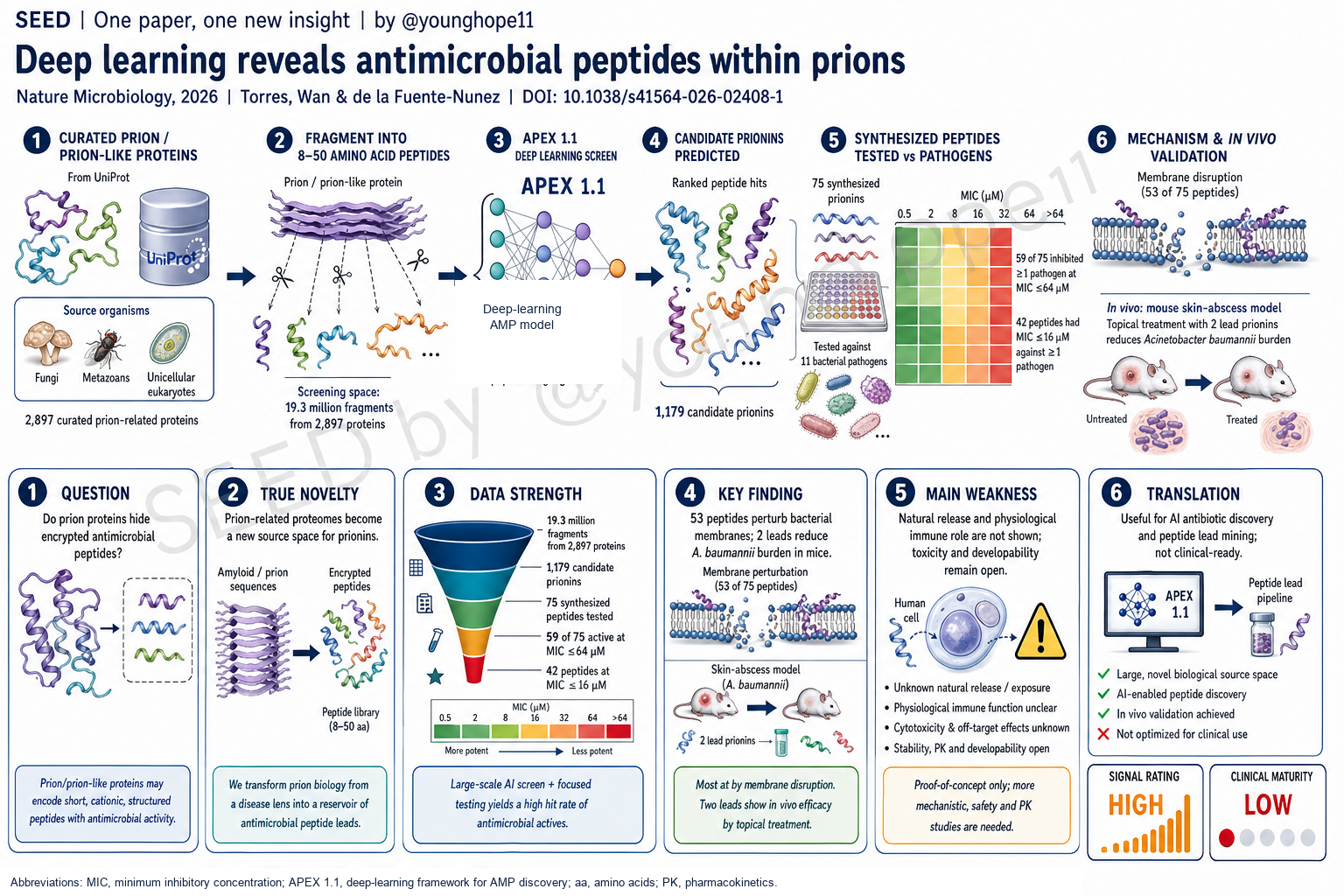
作者没有只从已知 AMP 数据库里找相似序列,而是从 UniProt 中 2,897 个被标注为 prion 或 prion-related 的条目出发,把每个蛋白切成所有 8-50 amino-acid fragments,得到 19,324,138 个 unique candidate peptides。随后用 APEX 1.1 预测它们对 11 个病原菌株的 MIC,并以 median predicted MIC <=64 µM 作为候选阈值,筛出 1,179 个候选 prionins。
这一步有两个关键点。第一,prionins 不是简单复刻已知 AMP。作者将它们与 DBAASP、APD3、DRAMP 和 in-house AMP training set 比较,发现 prionins 与已知 AMPs 只有部分重叠,整体上更长、更阳离子化、更疏水,但并不完全遵循传统 amphipathic AMP 模式。第二,候选 prionins 来自 1,068 个 source organisms,覆盖 fungi、metazoans 和 unicellular eukaryotes,说明这种潜在抗菌活性不是个别蛋白的孤例。
实验设计上,作者还对 75 个合成候选设置了多样性过滤:与已知 AMP 和入选候选之间都要求低于 70% sequence similarity。这让验证集更像是在测试一个新的 sequence class,而不只是挑选最像已知抗菌肽的序列。
因此,这篇文章的概念推进在于:AI 不只是加速已知抗菌肽优化,而是把一个此前主要被疾病语境定义的蛋白空间转化为可实验验证的抗菌肽来源。
数据强在哪里?
数据最强在“筛选规模大、实验命中率高、机制和体内模型都有闭环”。
第一,计算筛选规模足够大。作者从 2,897 个 prion-related proteins 中枚举出 19.3 million fragments,并用 APEX 1.1 对 11 个病原菌株做多任务 MIC 预测。APEX 1.1 的训练包括 1,642 个 in-house peptides、15,718 个 MIC measurements,以及 19,564 个 public AMPs 和 9,857 个 non-AMPs。这让模型不是只做 AMP/non-AMP classification,而是直接预测不同病原菌上的活性强弱。
第二,体外命中率很高。75 个合成 prionins 中,59 个在 <=64 µM 下抑制至少一个 bacterial pathogen,42 个在至少一个 pathogen 上达到 MIC <=16 µM。测试面板包括 A. baumannii、E. coli、K. pneumoniae、P. aeruginosa、S. aureus、MRSA、VRE E. faecalis、VRE E. faecium,以及 polymyxin/colistin-resistant E. coli AIC222。活性主要集中在 Gram-negative bacteria,这对耐药革兰阴性菌方向有实际意义。
第三,机制数据支持 membrane perturbation。作者用 circular dichroism 观察部分 prionins 在 membrane-mimetic 或 helix-promoting environments 中形成结构,再用 NPN uptake 测 outer-membrane permeabilization,用 DiSC3-5 测 cytoplasmic-membrane depolarization。总体上,53 个 prionins 产生膜扰动信号,提示膜破坏是这一类 peptide 的常见作用模式。
第四,选择性数据不是空白。人红细胞 haemolysis 很少,75 个 prionins 中只有 1 个在测试浓度范围内有 haemolytic activity。HEK293T cytotoxicity 更常见,说明不是所有 prionins 都可开发;但仍有 16 个 active peptides 在最高测试浓度下既没有可测 haemolysis,也没有可测 cytotoxicity。这为后续 lead prioritization 留出了空间。
第五,动物模型提供了早期体内验证。作者选择 prionin-7 和 prionin-38,分别来自 Thelonectria olida 和 Caenorhabditis elegans,在 6 周龄雌性 CD-1 mice 的 A. baumannii skin-abscess model 中测试。单次 topical treatment 在感染后 1 小时给药,剂量为 10x MIC,每组 n=6。第 2 天两者均显著降低 bacterial burden,效果接近 polymyxin B;第 4 天 prionin-38 仍维持约 3-log burden reduction,prionin-7 约 1-log reduction,且没有观察到治疗相关体重下降。
最大弱点是什么?
最大弱点是生理功能没有被证明。作者很清楚地指出,这项研究没有证明 prionins 会在感染时自然释放,也没有证明它们在原始宿主中作为 innate immune effectors 发挥作用。因此,“prion 里藏着抗菌肽”目前首先是 discovery source 的证据,不是 prion biology 中自然免疫功能的因果证明。
第二个弱点是候选空间依赖数据库标注和片段枚举。UniProt 的 prion/prion-related 条目包括 reviewed、unreviewed 和 isoform entries,标注质量和生物学语境并不完全一致。把蛋白切成 8-50 aa 片段是发现 encrypted peptides 的有效策略,但这些片段是否会被体内真实 proteolysis 生成,是另一个问题。
第三个弱点是机制仍偏初步。膜通透化和膜电位崩塌是 AMP 常见机制,但这不能排除其他靶点,也不能说明每个 prionin 都以同样方式杀菌。文章没有系统测试 resistance emergence、serum binding、salt sensitivity、protease stability、biofilm activity、post-antibiotic effect 或 host immunomodulation。
第四个弱点是体内验证仍是小规模局部感染模型。两个 lead 只在 A. baumannii skin abscess 中做 topical treatment,且 n=6 per group。这个模型适合证明早期 anti-infective activity,但不能直接外推到 systemic infection、肺部感染、血流感染或复杂组织环境。也还没有 pharmacokinetics、biodistribution、repeat dosing、局部刺激和长期安全数据。
第五个弱点是可开发性仍未优化。Peptide antibiotics 常见瓶颈包括体内稳定性、肾清除、生产成本、宿主毒性和给药途径。虽然 haemolysis 很少、部分 peptide 选择性较好,但 cytotoxicity 更常见,说明这个库还需要 medicinal chemistry、formulation 和 structure-activity optimization。
是否有临床转化意义?
有,但更像一个 antibiotic discovery platform signal,而不是马上能进入临床的候选药物。
第一,它拓展了抗菌肽发现的搜索空间。过去 de la Fuente-Nunez 团队已经从 human proteome、ancient proteomes、archaeal proteome 和 venoms 中挖掘 encrypted peptide antibiotics。这篇文章把同一逻辑推进到 prion/prion-like proteomes,说明 AI 可以系统地把“非免疫蛋白类别”转化为可实验验证的 antimicrobial lead libraries。
第二,它对耐药革兰阴性菌方向有现实意义。A. baumannii、P. aeruginosa、K. pneumoniae 和 polymyxin-resistant E. coli 都属于抗菌药开发中的高优先级问题。Prionins 的活性主要偏向 Gram-negative bacteria,再加上膜扰动机制和 A. baumannii 小鼠局部感染模型数据,使它们值得作为 topical、wound infection 或局部递送方向的早期 lead pool。
第三,它给 AI antibiotic discovery 提供了一个较完整的验证范式:大规模 sequence mining、相似性过滤、体外 MIC、膜机制、宿主细胞毒性、再到小鼠感染模型。这比单纯发布一批模型预测 peptide 更有说服力。
但临床距离仍远。下一步最关键的是对 prionin-7、prionin-38 和其他 selective leads 做序列优化、D-amino acid 或 cyclization 稳定化、serum/protease stability、PK/PD、resistance selection、biofilm testing、复杂感染模型和系统毒性评估。只有当这些问题解决后,prionins 才可能从“新来源空间”走向真实抗菌药候选。
Yang 的信号评级:High
理由:我会把这篇论文的科研信号评为 High。它的强点不是某一个 peptide 的临床前药效已经足够成熟,而是清楚证明 prion-related proteome 是一个意外丰富的 encrypted antimicrobial peptide source space。19.3 million fragments 的 AI 筛选、75 个 peptide 的合成验证、59 个体外活性、53 个膜扰动信号和 2 个小鼠感染模型 lead,让这个概念超过单纯计算预测。
这个 High 是“发现平台和概念信号 High”,不是“药物成熟度 High”。它把 prion biology、amyloidogenic sequences 和 antimicrobial peptide discovery 连接起来,也为 AI 挖掘非典型蛋白空间提供了可复制范式。
临床成熟度我会评为 Low。当前证据足以支持继续做 lead optimization 和感染模型扩展,但还不足以支持临床候选判断。真正的转化价值取决于 prionins 能否在稳定性、毒性、PK/PD、抗耐药演化和更复杂感染模型中保留优势。
Marcelo D. T. Torres, Fangping Wan and the team of Cesar de la Fuente-Nunez at the University of Pennsylvania Machine Biology Group recently reported a systematic deep-learning screen of prion and prion-like proteins using APEX 1.1. The study identifies a class of encrypted antimicrobial peptides, which the authors call prionins, and provides a new source space and validation path for mining antibiotic leads from unconventional protein classes.
What is the research question?
This paper asks a counterintuitive question: are prion and prion-like proteins only associated with misfolding, amyloid formation and neurodegenerative pathology, or can their primary sequences also hide antimicrobial peptides that could be released, cleaved or artificially mined?
The question has a real biological basis. Amyloid-beta, the cellular prion protein and other amyloid-associated proteins have been reported to show antimicrobial or host-protective activity. The authors connect that observation to encrypted peptide antibiotic discovery: if many non-immune proteins contain hidden antimicrobial fragments, then aggregation-prone and structurally unusual prion-related proteomes may represent an underexplored peptide mine.
More specifically, the study asks three things. First, do prion-related proteins contain large numbers of short fragments predicted by AI to be antimicrobial? Second, do these fragments actually inhibit clinically relevant bacteria in vitro and show an interpretable membrane-perturbing mechanism? Third, can selected lead peptides produce anti-infective activity in an animal infection model?
What is truly new?
The real novelty is the reframing of prions from a pathology-associated protein class into a sequence source space for antibiotic discovery.
The authors do not merely search known AMP databases for similar sequences. They start from 2,897 UniProt entries annotated as prion or prion-related, fragment each protein into all 8-50 amino-acid substrings, and obtain 19,324,138 unique candidate peptides. APEX 1.1 then predicts MIC values against 11 bacterial pathogens. Using a median predicted MIC threshold of <=64 uM, the authors identify 1,179 candidate prionins.
Two points matter. First, prionins are not simply copies of known AMPs. When compared with DBAASP, APD3, DRAMP and the in-house AMP training set, prionins overlap only partially with known antimicrobial peptides. On average, they are longer, more cationic and more hydrophobic, but they do not uniformly maximize classical amphipathic AMP patterns. Second, candidate prionins arise from 1,068 source organisms across fungi, metazoans and unicellular eukaryotes, suggesting that this potential activity is not a single-protein oddity.
The experimental set is also diversity-filtered. For the 75 synthesized candidates, the authors enforce less than 70% sequence similarity to known AMPs and to one another. That makes the validation closer to testing a new sequence class rather than simply choosing sequences most similar to known antimicrobial peptides.
The conceptual advance is therefore that AI is not only accelerating optimization of known AMPs. It is converting a protein space mostly defined by disease biology into an experimentally testable source of antimicrobial peptides.
Where is the data strongest?
The data are strongest because the study combines a large search space, a high experimental hit rate, mechanistic assays and an in vivo infection model.
First, the computational screen is large. The authors enumerate 19.3 million fragments from 2,897 prion-related proteins and use APEX 1.1 to perform multitask MIC prediction against 11 pathogenic strains. APEX 1.1 is trained on 1,642 in-house peptides with 15,718 MIC measurements, plus 19,564 public AMPs and 9,857 non-AMPs. The model is therefore not only classifying AMP versus non-AMP; it is estimating pathogen-specific antibacterial activity.
Second, the in vitro hit rate is high. Among 75 synthesized prionins, 59 inhibit at least one bacterial pathogen at <=64 uM, and 42 reach MIC <=16 uM against at least one pathogen. The test panel includes A. baumannii, E. coli, K. pneumoniae, P. aeruginosa, S. aureus, MRSA, VRE E. faecalis, VRE E. faecium and polymyxin/colistin-resistant E. coli AIC222. Activity is concentrated against Gram-negative bacteria, which is relevant for difficult antimicrobial development problems.
Third, the mechanistic data support membrane perturbation. The authors use circular dichroism to examine structure in membrane-mimetic or helix-promoting environments, NPN uptake to assess outer-membrane permeabilization, and DiSC3-5 to assess cytoplasmic-membrane depolarization. Overall, 53 prionins produce membrane-perturbing signals, supporting membrane disruption as a common mechanism for this class.
Fourth, selectivity is not ignored. Haemolysis against human red blood cells is rare: only 1 of 75 prionins shows haemolytic activity within the tested concentration range. HEK293T cytotoxicity is more frequent, so not every prionin is developable. Still, 16 active peptides show neither measurable haemolysis nor cytotoxicity at the highest concentrations tested, leaving room for lead prioritization.
Fifth, the animal data provide early in vivo validation. The authors select prionin-7 and prionin-38, derived from Thelonectria olida and Caenorhabditis elegans, respectively, and test them in a 6-week-old female CD-1 mouse A. baumannii skin-abscess model. A single topical treatment is administered 1 hour after infection at 10x MIC, with n=6 mice per group. At day 2, both peptides significantly reduce bacterial burden relative to untreated controls with efficacy comparable to polymyxin B. At day 4, prionin-38 maintains an approximately 3-log burden reduction, while prionin-7 shows a more modest approximately 1-log reduction. No treatment-associated weight loss is observed.
What is the biggest weakness?
The biggest weakness is that physiological function is not proven. The authors explicitly state that the study does not show that prionins are naturally released during infection or that they function as innate immune effectors in their native hosts. For now, the claim that prions hide antimicrobial peptides is strongest as discovery-source evidence, not as causal proof of a natural immune function in prion biology.
A second limitation is that the candidate space depends on database annotation and exhaustive fragmentation. The UniProt prion/prion-related entries include reviewed, unreviewed and isoform records, and the biological context of those annotations is not uniform. Cutting proteins into 8-50 amino-acid fragments is powerful for finding encrypted peptides, but whether those fragments are generated by real proteolysis in vivo is a separate question.
A third limitation is that the mechanism remains early-stage. Membrane permeabilization and membrane-potential collapse are common AMP mechanisms, but they do not exclude other targets and do not prove that every prionin kills through the same mechanism. The paper does not systematically test resistance emergence, serum binding, salt sensitivity, protease stability, biofilm activity, post-antibiotic effects or host immunomodulation.
A fourth limitation is that the in vivo validation is a small local infection model. The two leads are tested only as topical treatments in an A. baumannii skin abscess model, with n=6 mice per group. This is appropriate for early anti-infective validation, but it does not translate directly to systemic infection, lung infection, bloodstream infection or complex tissue environments. Pharmacokinetics, biodistribution, repeat dosing, local irritation and long-term safety are not yet established.
A fifth limitation is that developability is not optimized. Peptide antibiotics often face problems with stability, renal clearance, production cost, host toxicity and route of administration. Haemolysis is rare and some peptides look selective, but cytotoxicity is more frequent, so this library still needs medicinal chemistry, formulation and structure-activity optimization.
Is there translational or clinical relevance?
Yes, but the relevance is best understood as an antibiotic-discovery platform signal rather than a near-clinical drug-candidate signal.
First, the paper expands the search space for antimicrobial peptides. The de la Fuente-Nunez team has previously mined encrypted peptide antibiotics from human proteomes, ancient proteomes, archaeal proteomes and venoms. This paper pushes the same logic into prion and prion-like proteomes, showing that AI can turn non-immune protein categories into experimentally validated antimicrobial lead libraries.
Second, the work is relevant to resistant Gram-negative bacteria. A. baumannii, P. aeruginosa, K. pneumoniae and polymyxin-resistant E. coli are high-priority antimicrobial challenges. The fact that prionin activity is concentrated against Gram-negative bacteria, together with membrane perturbation and A. baumannii mouse skin-infection data, makes these peptides worth exploring as early leads for topical, wound-infection or local-delivery applications.
Third, the study provides a relatively complete validation template for AI antibiotic discovery: large-scale sequence mining, similarity filtering, in vitro MIC testing, membrane-mechanism assays, host-cell toxicity testing and a mouse infection model. That is more persuasive than releasing a list of model-predicted peptides alone.
Clinical distance remains substantial. The next decisive work is sequence optimization of prionin-7, prionin-38 and other selective leads, including D-amino acid substitution or cyclization for stability, serum and protease stability, PK/PD, resistance selection, biofilm testing, more complex infection models and systemic toxicity. Only if those steps succeed can prionins move from a new source space toward true antimicrobial drug candidates.
Yang’s signal rating: High
Reason: I would rate the scientific signal of this paper as High. The strongest point is not that any one peptide is already a mature preclinical drug candidate, but that the study clearly establishes prion-related proteomes as an unexpectedly rich source space for encrypted antimicrobial peptides. The combination of a 19.3-million-fragment AI screen, 75 synthesized peptides, 59 in vitro actives, 53 membrane-perturbing peptides and 2 mouse infection-model leads moves the concept beyond computational prediction.
This High means discovery-platform and conceptual signal High, not drug-maturity High. The paper connects prion biology, amyloidogenic sequence space and antimicrobial peptide discovery, while providing a reproducible template for AI-guided mining of unconventional protein spaces.
I would rate clinical maturity as Low. The evidence is strong enough to justify lead optimization and broader infection-model testing, but not enough to support clinical-candidate judgment. The real translational value will depend on whether prionins can retain advantages after optimization for stability, toxicity, PK/PD, resistance emergence and more complex infection models.