SEED | 基因编辑安全性,不能只按平台下结论 SEED | Gene-editing safety cannot be judged by platform alone AI-assisted · reviewed
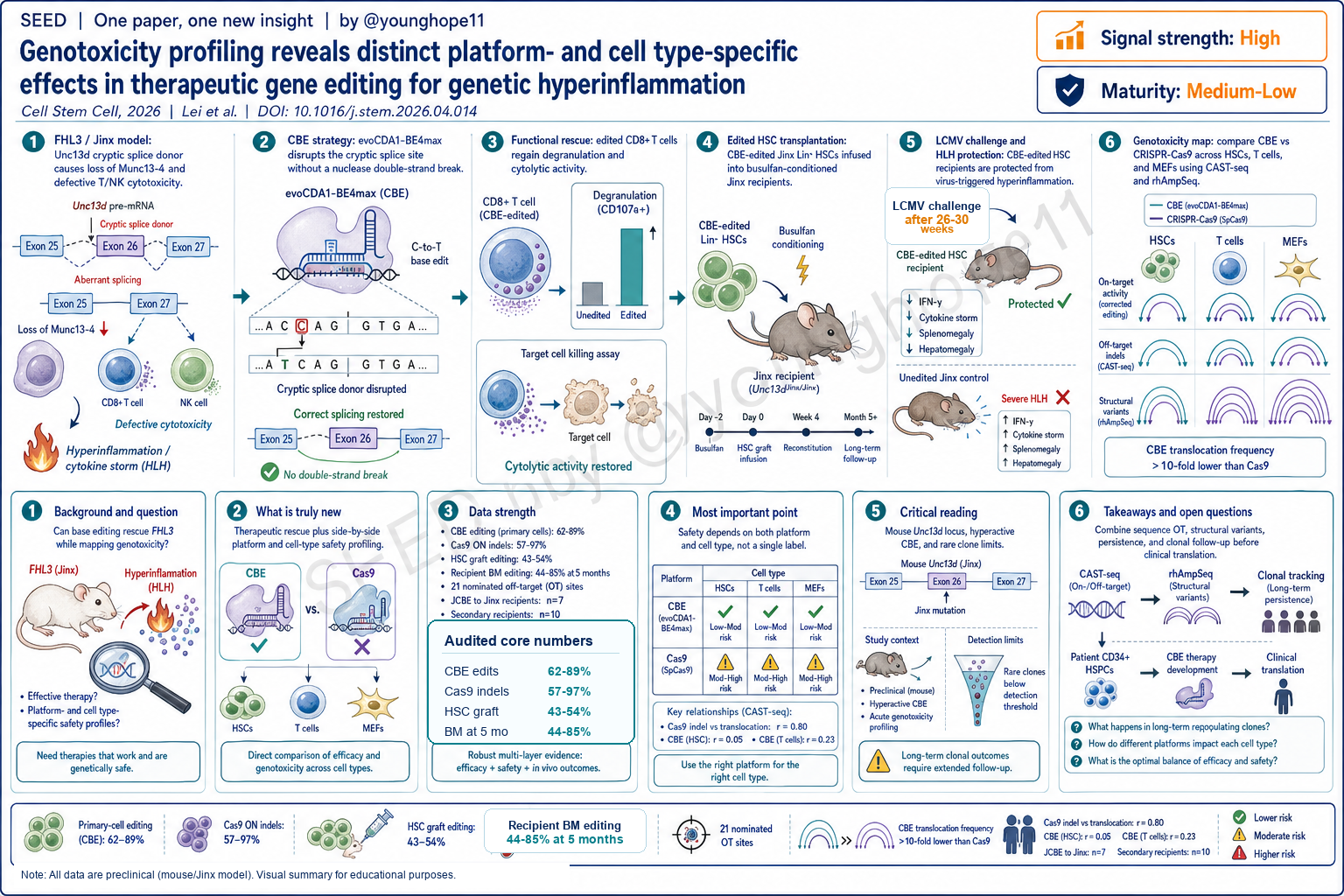
University of Freiburg 的 Lei Lei 与 Toni Cathomen 团队近期报道,cytosine base editing 可以纠正 FHL3/Jinx 小鼠 Unc13d 剪接缺陷并保护受体小鼠免于 LCMV 触发的 HLH 样高炎症,同时通过 CAST-seq 与 rhAmpSeq 对 CBE 和 CRISPR-Cas9 在 HSC、T 细胞和 MEF 中的基因毒性进行并排比较,为基因编辑疗法的细胞类型特异性安全评估提供了新的框架。
FHL3 给了 base editing 一个真实的安全测试场景
这篇论文研究的是 familial hemophagocytic lymphohistiocytosis type 3。FHL3 由 UNC13D/Munc13-4 功能缺陷导致,T 细胞和 NK 细胞杀伤功能受损,抗原刺激后容易进入持续免疫激活、IFN-gamma 升高、巨噬细胞活化和多器官损伤。
模型是 Jinx 小鼠。它在 Unc13d intron 26 中有一个 C-to-A 变异,形成 cryptic splice donor,导致异常剪接和 Munc13-4 缺失。作者的问题不是单纯证明 base editing 能改一个碱基,而是问:能否用 CBE 恢复细胞毒免疫,并且在同一套实验中把 CBE 与 CRISPR-Cas9 的 off-target 和结构变异风险做细胞类型分层比较?
新意在于把疗效救援和基因毒性图谱放在一起
作者先在 Jinx reporter cell line 中筛选多个 base editor,最后选择 evoCDA1-BE4max 作为高活性的 CBE。随后在 Jinx CD8+ T 细胞中编辑 Unc13d cryptic splice site,恢复 exon 26 到 exon 27 的正常剪接,并恢复 CD107a degranulation 和抗原特异性 cytolytic activity。
更重要的是,文章没有停留在 T 细胞体外功能。作者编辑 Jinx Lin- HSC,移植到 busulfan-conditioned Jinx recipient,然后在 26-30 周后用 LCMV 触发 HLH。CBE-edited HSC 受体在感染后维持体重、降低 hepatosplenomegaly 和 IFN-gamma,T 细胞 cytotoxicity 恢复,并清除病毒触发因素。
真正的新意是第二层:同一靶点、同一 gRNA、三类原代细胞中并排比较 CBE 和 CRISPR-Cas9。安全结论不是“CBE 更安全”或“Cas9 更危险”这种单轴判断,而是平台、细胞类型和时间共同决定基因毒性图谱。
数据强在三层:功能救援、CAST-seq 和长期追踪
第一层是疗效。CBE 在 primary murine CD8+ T cells 中达到 71% target cytosine conversion;在移植前 Lin- HSC 中 Unc13d-edited allele 为 43%-54%。Jinx recipient 长期随访到 28-32 周时,编辑比例在骨髓中为 33%-84%,在脾脏中为 32%-78%,并显示多谱系植入。
第二层是平台和细胞类型比较。作者在 Lin- HSC、CD8+ T cell 和 mouse embryonic fibroblast 中比较 CBE 与 CRISPR-Cas9。on-target 编辑都很高:Cas9 indels 为 57%-97%,CBE base substitutions 为 62%-89%。CAST-seq 显示两种平台、三种细胞都可能产生 on-target aberrations 和 off-target-mediated translocations,但模式不同:Cas9 结构变异更强,尤其在 HSC 中;CBE 总 translocation frequency 比 Cas9 低超过 10 倍,但 off-target sequence editing 更广、更难用单一指标预测。
第三层是长期。5 个月后,7 只接受 CBE-edited HSC 的 Jinx recipient 骨髓中仍可检测到 44%-85% 的 Unc13d base editing;off-target base substitutions 持续存在,但 CAST-seq 未检测到 chromosomal translocations,secondary transplantation 也没有显示 base-editing-associated leukemogenesis。
最重要的一点:安全性是平台加细胞类型的函数
这篇文章最值得记住的不是某个单一百分比,而是安全评估逻辑的改变。Cas9 的 indel frequency 与 translocation burden 有较强相关性,相关系数 r=0.80;但在 CBE 中,CAST-seq hits 与 indels 的相关性只有 r=0.05,与 base substitutions 的相关性为 r=0.23。
这意味着,不能用一个 amplicon editing percentage 或一个 off-target indel readout 来替代结构变异风险判断。CBE 的 sequence-level off-target 可以很广,但不总是变成 translocation;反过来,低频位点也可能参与结构变异。HSC、T 细胞和 fibroblast 的 DNA repair、cell cycle、chromatin accessibility 和选择压力不同,最后留下的安全图谱也不同。
批判性阅读:这是强框架,但不是临床安全结论
第一,模型和靶点仍然集中在一个 murine Unc13d locus。作者也测试了额外 guide,但核心疗效和长期追踪仍围绕 Jinx/FHL3 模型,不能直接外推到所有 base editing 药物。
第二,使用的是高活性的 evoCDA1-BE4max。它让 murine primary cells 达到足够编辑效率,但也可能放大 CBE 相关 indels 和 off-target base substitutions。临床上不同 editor、delivery modality、exposure time 和细胞状态都会改变风险。
第三,HSC 来自 disease manifestation 前的健康小鼠,而真实 FHL3 患者 HSC 可能经历炎症压力。in vivo 追踪还存在 oligoclonal repopulation 和 sampling depth 限制,不能排除非常稀有的异常 clone。一个 secondary recipient 出现 lymphoproliferative/leukemia-like disease,但作者判断为 host-origin、与 busulfan conditioning 更相关,这也提醒我们长期归因不能只看编辑工具本身。
对基因编辑疗法的启发
这篇文章对未来药物开发的启发很直接:base editing 的安全包不能只做预测 off-target 或几个 amplicon 位点。至少在 HSC 和免疫细胞疗法中,需要把 sequence-level OT、large on-target aberration、off-target-mediated translocation、cell-type persistence 和长期 clonal behavior 放在同一个风险框架里。
对 FHL3 来说,文章也提供了一个可继续发展的治疗路径。FHL mutation database 中 183 个 UNC13D variants 有定义的致病机制,其中 138 个是 single-nucleotide substitutions,95 个理论上属于当前 CBE/ABE 可处理的 transition mutations。但真正走向患者,需要 mutation-tailored guide/editor 设计,以及在患者 CD34+ cell workflow 中重新做细胞类型特异性安全评估。
Yang 的信号评级:High
轴一,信号强度:High。理由:这篇文章同时提供了功能性疾病救援和系统性基因毒性图谱,不只是证明 CBE 能治疗一个小鼠模型,而是把“平台效应”和“细胞类型效应”变成基因编辑安全评估的核心变量。
轴二,成熟度:Medium-Low。理由:治疗证据仍是小鼠模型,且集中在单一 locus、特定 CBE 和特定 delivery 条件;对人 CD34+ 细胞、不同 editor 和真实患者炎症背景的可转化安全性仍需验证。
一句话总结:这篇论文提醒我们,基因编辑安全性不是某个平台的固定属性,而是 editor、靶点、细胞类型和时间共同塑造出来的结果。
The Lei Lei and Toni Cathomen team at the University of Freiburg recently reported that cytosine base editing can correct the Unc13d splicing defect in an FHL3/Jinx mouse model and protect recipient mice from LCMV-triggered HLH-like hyperinflammation, while side-by-side CAST-seq and rhAmpSeq profiling of CBE and CRISPR-Cas9 across HSCs, T cells, and MEFs provides a new framework for cell type-specific safety assessment of therapeutic genome editing.
FHL3 makes base-editing safety concrete
The disease context is familial hemophagocytic lymphohistiocytosis type 3. FHL3 is caused by UNC13D/Munc13-4 dysfunction, which impairs T and NK cell cytotoxicity and can drive sustained immune activation, high IFN-gamma, macrophage activation, and multi-organ injury.
The model is the Jinx mouse. It carries a C-to-A variant in Unc13d intron 26 that creates a cryptic splice donor, causing aberrant splicing and loss of Munc13-4. The paper therefore asks more than whether base editing can fix one base. It asks whether CBE can restore cytotoxic immunity while the same experimental system compares CBE and CRISPR-Cas9 off-target and structural-variant risks across cell types.
The novelty is linking rescue with a genotoxicity map
The authors first screened base editors in a Jinx reporter cell line and selected evoCDA1-BE4max as the high-activity CBE. They then edited the Unc13d cryptic splice site in Jinx CD8+ T cells, restored normal exon 26 to exon 27 splicing, and rescued CD107a degranulation and antigen-specific cytolytic activity.
The paper does not stop at ex vivo T cell function. The authors edited Jinx Lin- HSCs, transplanted them into busulfan-conditioned Jinx recipients, and triggered HLH with LCMV 26 to 30 weeks later. Recipients of CBE-edited HSCs maintained body weight, had reduced hepatosplenomegaly and IFN-gamma, restored T cell cytotoxicity, and cleared the viral trigger.
The second layer is the real novelty: the same target, same gRNA, and three primary cell types were used to compare CBE and CRISPR-Cas9 side by side. The safety message is not a one-axis claim that CBE is safer or Cas9 is riskier. Platform, cell type, and time jointly shape the genotoxicity profile.
The strongest data combine rescue, CAST-seq, and follow-up
The first layer is therapeutic rescue. CBE achieved 71% target cytosine conversion in primary murine CD8+ T cells. In Lin- HSCs before transplantation, Unc13d-edited alleles were 43%-54%. At 28-32 weeks, edited cells ranged from 33%-84% in bone marrow and 32%-78% in spleen, with multilineage reconstitution.
The second layer is platform and cell type comparison. The authors compared CBE and CRISPR-Cas9 in Lin- HSCs, CD8+ T cells, and mouse embryonic fibroblasts. On-target editing was high across the system: Cas9 indels ranged from 57%-97%, and CBE base substitutions ranged from 62%-89%. CAST-seq showed that both platforms and all three cell types can generate on-target aberrations and off-target-mediated translocations. The patterns differed: Cas9 produced stronger structural rearrangement signals, especially in HSCs; CBE had more than 10-fold lower total translocation frequency than Cas9, but a broader and less predictable sequence-level off-target footprint.
The third layer is long-term tracking. Five months after transplantation, bone marrow from seven Jinx recipients still carried 44%-85% Unc13d base editing. Off-target base substitutions persisted, but CAST-seq detected no chromosomal translocations, and secondary transplantation did not reveal base-editing-associated leukemogenesis.
The main lesson is that safety is platform plus cell type
The key point is not a single editing percentage. It is the safety logic. For Cas9, indel frequency correlated strongly with translocation burden, with r=0.80. For CBE, CAST-seq hits correlated poorly with indels (r=0.05) and base substitutions (r=0.23).
This means that an amplicon editing percentage or a single off-target indel readout cannot stand in for structural-variant risk. CBE sequence-level off-target activity can be broad without always becoming translocations; conversely, lower-frequency sites can still contribute to structural events. HSCs, T cells, and fibroblasts differ in DNA repair, cell cycle, chromatin accessibility, and selection pressure, and the safety footprint left behind is correspondingly different.
Critical reading: a strong framework, not a clinical safety answer
First, the model and target remain centered on one murine Unc13d locus. The authors tested additional guides, but the core rescue and long-term follow-up still revolve around the Jinx/FHL3 model and cannot be directly generalized to all base-editing therapies.
Second, the study uses the highly active evoCDA1-BE4max. That activity made editing possible in murine primary cells, but it may also amplify CBE-associated indels and off-target base substitutions. In the clinic, editor choice, delivery modality, exposure time, and cell state will all change risk.
Third, the HSCs came from healthy mice before disease manifestation. Real FHL3 patient HSCs may experience inflammatory stress. In vivo tracking is also limited by oligoclonal repopulation and sampling depth, so very rare abnormal clones cannot be excluded. One secondary recipient developed lymphoproliferative or leukemia-like disease, but the authors attributed it to host origin and busulfan conditioning rather than the edited graft, which still illustrates how carefully adverse outcomes must be assigned.
What this should change for genome-editing translation
The translational message is direct: a base-editing safety package should not rely only on predicted off-target sites or a few amplicon assays. At least for HSC and immune-cell therapies, sequence-level off-target events, large on-target aberrations, off-target-mediated translocations, cell type-specific persistence, and long-term clonal behavior need to be interpreted together.
For FHL3, the paper also outlines a plausible development path. The FHL mutation database lists 183 UNC13D variants with defined pathogenic mechanisms; 138 are single-nucleotide substitutions, and 95 are in principle transition mutations addressable by current CBEs or ABEs. But translation would require mutation-tailored guide/editor design and renewed cell type-specific safety profiling in a patient CD34+ cell workflow.
Yang’s signal rating: High
Signal strength: High. The paper combines functional disease rescue with systematic genotoxicity profiling. It is not only a demonstration that CBE can treat a mouse model; it turns platform effects and cell type effects into core variables for genome-editing safety assessment.
Maturity: Medium-Low. The therapeutic evidence remains a mouse-model result centered on one locus, one CBE, and one delivery setting. Human CD34+ cell translation, editor generalizability, and safety in an inflammatory patient context remain to be tested.
One-sentence summary: Genome-editing safety is not a fixed property of a platform; it is shaped by editor, target, cell type, and time.