SEED | 碱基编辑揭示 NANOG 的人类胚胎角色 SEED | Base editing reveals NANOG's human embryo role AI-assisted · reviewed
University of Cambridge 与 Francis Crick Institute 等机构的 Oliver J. Bower 与通讯作者 Kathy K. Niakan、Katarina Harasimov 团队近期在 Nature 报道,ABE8e 碱基编辑可在人类胚胎中无双链断裂地破坏 NANOG 剪接供体,从而揭示 NANOG 对人类上胚层指定和多能性维持必不可少,并显示人类 primitive endoderm 命运与小鼠模型存在关键差异。
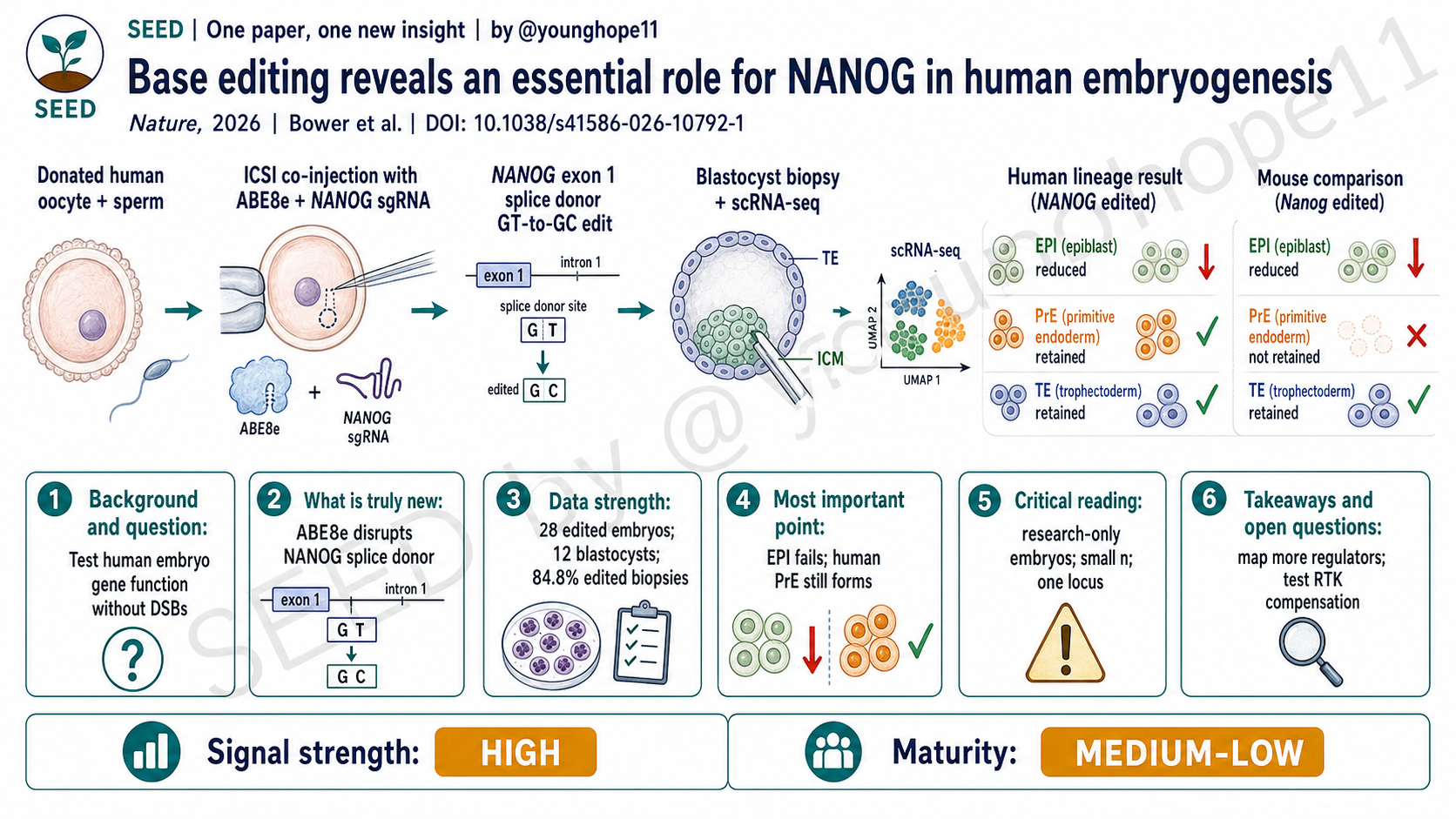
为什么要在人类胚胎中直接测试 NANOG
NANOG 是早期胚胎发育和多能性网络里的核心转录因子。在小鼠中,Nanog 对 epiblast 形成很重要;在人类胚胎中,NANOG 表达也高度集中在 epiblast,也就是未来 embryo proper 的前体细胞群。但“表达在这里”不等于“功能一定相同”。人类早期胚胎研究长期受伦理、样本稀缺和技术限制影响,很多关键调控因子只能从小鼠或 human ESC 间接推断。
这篇文章问的是一个很具体的问题:NANOG 在人类 preimplantation embryo 里到底是不是 epiblast specification 必需因子?如果是,它对 primitive endoderm 和 trophectoderm 的影响是否和小鼠一样?
过去用 CRISPR-Cas9 在人类胚胎中做功能遗传学,有一个大问题:双链 DNA 断裂可能造成大片段缺失、染色体重排和非整倍体。人类合子对 DSB repair 尤其敏感,这会让研究者难以分辨观察到的表型究竟来自目标基因缺失,还是来自编辑本身造成的基因组损伤。因此,这篇文章的背景不是“能不能编辑胚胎”这么简单,而是“能不能更干净地在人类胚胎中测试基因功能”。
新意是用 ABE8e 做无 DSB 的功能扰动
作者没有用 Cas9 nuclease 直接敲除 NANOG,而是用 adenine base editor ABE8e 靶向 NANOG exon 1 的 splice donor site,把 canonical GT splice donor 改成 GC,诱导剪接缺陷和功能性 loss-of-function。这个设计的价值在于,它绕开了 DSB,同时通过剪接位点改变实现基因功能扰动。
研究流程先在人类 ESC 中优化 sgRNA 和编辑策略。NANOG splice donor 编辑在 primed human ESC 中导致 NANOG 蛋白丢失、多能性标志下调、神经外胚层相关基因上调;单细胞克隆实验中,AAVS1 对照克隆 95.8% 能形成稳定克隆,而 NANOG 编辑克隆第 7 天只有 41.7% 形成克隆,第 14 天无一持续。这说明 NANOG 对 human ESC 自我更新非常关键,也为胚胎实验提供了功能验证。
真正进入胚胎实验时,作者用 ICSI 共注射 sperm、ABE8e mRNA 或蛋白、以及 NANOG sgRNA。28 个 NANOG-edited human embryos 中,12 个形成 blastocyst,比例为 43%。随后,作者对 blastocyst 做免疫染色、trophectoderm biopsy、whole-genome sequencing、amplicon sequencing 和 polar region single-cell RNA-seq。这个设计把编辑结果、基因组安全性和细胞命运读数连在了一起。
所以,这篇文章的新意有两层:一是把 base editing 用作人类胚胎功能遗传学工具;二是用它直接回答 NANOG 在人类胚胎谱系指定中的作用,而不是继续依赖小鼠外推。
强数据来自编辑效率、安全性读数和单细胞谱系
第一组强数据是编辑效率。作者在 12 个 NANOG-edited blastocysts 的 46 个 trophectoderm biopsies 中检测到 84.8% 的 on-target T•A>C•G 编辑,具体为 39/46 biopsies。biallelic editing 在 biopsy 间存在差异:37.0% 为 homozygous mutation,47.8% 为 heterozygous 或 mosaic mutation。bystander edit 也很常见,12 个 blastocysts 中 10 个出现相邻 silent bystander edit,但这个改变不改变 NANOG 氨基酸序列。
第二组强数据是 genotoxicity 和 off-target 读数。作者对 23 个 trophectoderm biopsies 做 low-pass whole-genome sequencing,没有在 on-target locus 或 top 5 predicted off-target sites 检测到染色体材料丢失。对 10 个 predicted off-target sites 做 amplicon sequencing 时,在 10 个 edited embryos 的 30 个 biopsies 中观察到的编辑率均不超过 0.15%。在 human ESC 的 RNA-seq off-target 分析中,ABE8e NANOG-edited 细胞 A-to-G 变化相对对照增加 1.2%,但其他非 ABE 方向的碱基变化也有类似量级,提示相当一部分可能来自技术或测序噪声。
第三组强数据是单细胞谱系。作者从 12 个 NANOG-edited blastocysts 中获得 11 个可用于 scRNA-seq 的 polar region 样本,共 359 个细胞,并与 2 个 AAVS1-edited control embryos 的 42 个细胞以及 888 个已发表人类 preimplantation embryo reference cells 整合分析。结果显示,3 个 homozygous NANOG mutant embryos 的细胞主要落在 ICM、trophectoderm 和 primitive endoderm 相关群体,而不是正常 epiblast 群体;epiblast markers 如 NANOG、LEFTY1、LEFTY2 和 FGF4 显著降低。
第四组数据是人鼠对照。小鼠 Nanog 扰动同样破坏 epiblast specification,但小鼠 Nanog-edited embryos 未能建立离散的 epiblast 或 primitive endoderm cells;相比之下,人类 NANOG-edited embryos 虽然 epiblast 失败,却仍保留 primitive endoderm 细胞,并表达 GATA6、GATA4、PDGFRA、SOX17、FOXA2、OTX2 以及 basement membrane 相关基因。这让表型从“一个基因敲除结果”变成了“人类发育调控不能简单复刻小鼠模型”的证据。
关键发现是 EPI 失败,但人类 PrE 仍然保留
这篇文章最重要的一点,是把 NANOG 的功能拆成了两个层面。
第一层是保守的:NANOG 对 epiblast specification 必不可少。无论在人类还是小鼠,NANOG/Nanog loss 都会破坏 pluripotent epiblast 的建立。这一点符合 NANOG 在多能性网络中的核心位置。
第二层是不保守的:primitive endoderm 的命运在人类和小鼠不同。小鼠中 Nanog loss 伴随 Fgf4 下调,primitive endoderm specification 失败;但在人类 NANOG-edited embryos 中,即使 FGF4 transcript 缺失,primitive endoderm 仍可保留。作者进一步看到 FGFR1-4、INSR、IGF1R,以及 FGF1、FGF12、FGF18、IGF1 等替代 RTK signalling 组件表达,提示人类胚胎中可能存在绕过 FGF4 的补偿机制。
这个点很重要。它说明,人类胚胎不是小鼠胚胎的缩小版,也不是 human ESC 体外状态的简单投影。某个转录因子在物种间可能保留一个核心功能,同时在相邻谱系或信号补偿上发生分化。对发育生物学来说,这类“部分保守、部分分叉”的结论比单纯说某个基因 essential 更有价值。
不能把它读成胚胎基因编辑临床路线
这篇文章必须严肃地区分“科研功能扰动工具”和“临床胚胎编辑路线”。
第一,研究使用的是捐赠给科研的人类 gametes 和 embryos,并在英国 HFEA 许可和伦理审批框架下完成。它的目标是理解早期人类发育,不是建立可临床使用的 heritable genome editing 流程。
第二,样本量仍然很小,这是人类胚胎研究不可避免但必须正视的限制。28 个 NANOG-edited embryos、12 个 blastocysts 和 11 个 scRNA-seq 样本足以提供强科学信号,但不足以定义 base editing 在人类胚胎中的完整安全谱。
第三,trophectoderm biopsy 的 genotype 不能完全代表整个 embryo。作者也明确指出,biopsy 只反映胚胎的一部分,可能不能完全对应 polar region 或 ICM 内每个细胞的真实 genotype。mosaicism 仍然是解释人类胚胎编辑实验时最需要谨慎的变量之一。
第四,安全性结论还不能泛化。这里看到 no obvious genotoxicity 和 low off-target editing,是在一个位点、一个编辑策略、有限样本和特定检测深度下得到的。不同基因、不同 sgRNA、不同编辑器、不同递送方式,风险可能完全不同。作者也强调,未来必须在更多 genomic loci、更大 cohort 和更全面的 safety assays 中验证。
第五,论文中有 genome editing 领域重要利益相关方参与,包括 David R. Liu 与 base editing/prime editing 相关公司的关系和专利利益披露。这不削弱数据本身,但提醒读者:平台安全性和可推广性的判断必须依赖独立重复和更广泛评估。
下一步要把补偿机制和平台边界做实
这项研究最直接的下一步,是找出人类 NANOG loss 后 primitive endoderm 为什么还能保留。FGF18、IGF1 或其他 RTK signalling 组件是否真的补偿了 FGF4?这是表达相关,还是功能必需?需要通过更精确的扰动组合来验证。
第二个方向是扩展发育调控因子。NANOG 是一个高价值起点,但真正改变人类胚胎功能遗传学的,是能否系统测试 OCT4、SOX2、GATA6、TFAP2C、CDX2 等关键节点,并区分哪些功能在小鼠、人类和 stem-cell models 之间保守,哪些发生物种特异性重排。
第三个方向是平台安全性。ABE8e 这次比 Cas9 nuclease 更少看到明显 genotoxicity,但这不是“安全”的终点,而是一个更好起点。需要更高分辨率的 on-target structural variant 检测、全基因组 off-target profiling、RNA off-target 分析、长期 developmental potential 评估,以及不同编辑器变体在 human embryos 中的直接比较。
第四个方向是伦理和治理。即使 base editing 能让人类胚胎功能研究更精确,临床 heritable genome editing 也不能由技术可行性自动推出。论文最后也明确强调,任何临床转化都必须以广泛社会讨论、严格伦理审查和公共支持为前提。
Yang 的信号评级:High
轴一,信号强度:High。 理由是它同时在技术和生物学两端给出强信号:技术上,ABE8e 为人类胚胎基因功能扰动提供了一个比 DSB nuclease 更干净的路径;生物学上,它直接证明 NANOG 对人类 epiblast specification 必需,并揭示 primitive endoderm 命运存在人鼠差异。
轴二,成熟度:Medium-Low。 理由是这仍是基础研究和平台验证阶段,样本稀缺、位点单一、mosaicism 和安全性泛化都没有解决;它适合推动人类发育功能遗传学,而不是支持临床胚胎编辑。
一句话总结:这篇文章最有价值的地方,是用碱基编辑把人类胚胎发育从“看见表达”推进到“直接测试功能”,同时提醒我们小鼠模型无法替代人类胚胎本身。
Oliver J. Bower, corresponding authors Kathy K. Niakan and Katarina Harasimov, and colleagues from the University of Cambridge, the Francis Crick Institute and collaborating centers recently reported in Nature that ABE8e base editing can disrupt the NANOG splice donor in human embryos without inducing double-strand breaks, revealing that NANOG is essential for human epiblast specification and pluripotency maintenance while exposing a key difference between human and mouse primitive endoderm fate.
Why NANOG has to be tested directly in human embryos
NANOG is a core transcription factor in early embryo development and pluripotency networks. In mouse embryos, Nanog is important for epiblast development. In human embryos, NANOG expression is also concentrated in the epiblast, the precursor of the embryo proper. But expression is not the same as function. Human embryo research has long been constrained by ethics, sample scarcity and technical limitations, so many key regulatory factors have been inferred indirectly from mouse embryos or human ESCs.
This paper asks a specific question: is NANOG truly required for epiblast specification in human preimplantation embryos, and does its effect on primitive endoderm and trophectoderm match the mouse model?
Earlier CRISPR-Cas9 functional genetics in human embryos faced a major problem: double-strand DNA breaks can create large deletions, chromosomal rearrangements and aneuploidy. Human zygotes are particularly vulnerable to inefficient DSB repair, making it difficult to know whether a phenotype comes from loss of the target gene or from editing-induced genomic damage. The background question is therefore not simply whether embryos can be edited, but whether gene function can be tested more cleanly in human embryos.
The real novelty is DSB-sparing functional genetics
The authors did not use Cas9 nuclease to knock out NANOG. Instead, they used the adenine base editor ABE8e to target the splice donor site in NANOG exon 1, changing the canonical GT splice donor to GC and inducing a splicing defect that functions as a loss-of-function perturbation. The value of this design is that it avoids DSBs while still disrupting gene function through splice-site editing.
The strategy was first optimized in human ESCs. NANOG splice-donor editing in primed human ESCs caused loss of NANOG protein, reduced pluripotency markers and increased neuroectoderm-associated transcripts. In single-cell colony cloning, 95.8% of AAVS1-edited control clones formed viable stable colonies, whereas only 41.7% of NANOG-edited clones formed colonies by day 7 and none persisted by day 14. This established NANOG as critical for human ESC self-renewal and provided a functional rationale for embryo experiments.
In the embryo experiments, the authors used ICSI to co-inject sperm, ABE8e mRNA or protein, and the NANOG sgRNA. Of 28 NANOG-edited human embryos, 12 formed blastocysts, or 43%. The authors then analyzed blastocysts by immunofluorescence, trophectoderm biopsy, whole-genome sequencing, amplicon sequencing and polar-region single-cell RNA-seq. This connected editing genotype, genome-safety readouts and cell-fate outcomes in one workflow.
The novelty is therefore twofold: using base editing as a tool for human embryo functional genetics, and using that tool to directly test NANOG’s role in human lineage specification rather than continuing to extrapolate from mouse models.
The strongest data link editing, genome safety and single-cell lineage state
The first strong dataset is editing efficiency. Across 46 trophectoderm biopsies from 12 NANOG-edited blastocysts, the authors detected on-target T-A to C-G editing in 84.8%, or 39 of 46 biopsies. Biallelic editing varied among biopsies: 37.0% showed homozygous mutation and 47.8% showed heterozygous or mosaic mutation. Bystander editing was also common, appearing in 10 of 12 blastocysts, but the bystander change was silent and did not alter the NANOG amino acid sequence.
The second strong dataset is genotoxicity and off-target assessment. Low-pass whole-genome sequencing of 23 trophectoderm biopsies detected no chromosomal material loss at the on-target locus or top five predicted off-target sites. Amplicon sequencing of 10 predicted off-target sites in 30 biopsies from 10 edited embryos found editing rates of 0.15% or less. In RNA-seq off-target analysis in human ESCs, ABE8e NANOG-edited cells had a 1.2% increase in A-to-G changes compared with controls, but non-ABE base changes appeared at similar scale, suggesting that much of this signal may reflect technical or sequencing noise.
The third strong dataset is single-cell lineage analysis. The authors obtained polar-region scRNA-seq data from 11 of 12 NANOG-edited blastocysts, yielding 359 cells, and integrated these with 42 cells from two AAVS1-edited control embryos and 888 published human preimplantation reference cells. Cells from three homozygous NANOG mutant embryos clustered mainly with ICM, trophectoderm and primitive endoderm groups rather than the normal epiblast group; epiblast markers including NANOG, LEFTY1, LEFTY2 and FGF4 were significantly reduced.
The fourth layer is species comparison. Mouse Nanog perturbation also disrupted epiblast specification, but mouse Nanog-edited embryos failed to establish discrete epiblast or primitive endoderm cells. In contrast, human NANOG-edited embryos failed to form epiblast properly yet retained primitive endoderm cells expressing markers such as GATA6, GATA4, PDGFRA, SOX17, FOXA2 and OTX2. That turns the finding from a single gene-perturbation result into evidence that human developmental regulation cannot be treated as a simple copy of the mouse model.
The core message is EPI failure with human PrE retention
The most important point is that the paper separates NANOG function into two layers.
The first layer is conserved: NANOG is essential for epiblast specification. In both human and mouse embryos, NANOG/Nanog loss disrupts establishment of the pluripotent epiblast. That fits NANOG’s central position in the pluripotency network.
The second layer is not conserved: primitive endoderm behaves differently between species. In mouse embryos, Nanog loss reduces Fgf4 and primitive endoderm specification fails. In human NANOG-edited embryos, primitive endoderm can still persist despite the absence of FGF4 transcripts. The authors observed expression of alternative RTK-signalling components, including FGFR1-4, INSR, IGF1R and ligands such as FGF1, FGF12, FGF18 and IGF1, suggesting a possible compensatory route around FGF4.
That distinction matters. Human embryos are not simply scaled-down mouse embryos, nor are they direct projections of human ESC culture states. A transcription factor may conserve one core function while diverging in adjacent lineage decisions or signalling compensation. For developmental biology, this kind of partial conservation and partial divergence is more informative than simply declaring a gene essential.
This is not a clinical germline-editing roadmap
The paper needs to be read as a basic functional-genetics study, not as a clinical embryo-editing roadmap.
First, the work used human gametes and embryos donated for research under UK HFEA licensing and ethics approval. The purpose was to understand early human development, not to establish a clinically usable heritable genome-editing workflow.
Second, the sample size remains small, as is unavoidable but important in human embryo research. Twenty-eight edited embryos, 12 blastocysts and 11 scRNA-seq samples are enough to generate a strong scientific signal, but not enough to define the full safety profile of base editing in human embryos.
Third, the genotype of a trophectoderm biopsy does not fully represent the entire embryo. The authors explicitly note that each biopsy samples only part of the embryo and may not match every cell in the polar region or ICM. Mosaicism remains one of the most important variables when interpreting human embryo editing experiments.
Fourth, the safety conclusions are not generalizable yet. The observed lack of obvious genotoxicity and low off-target editing apply to one locus, one editing strategy, limited samples and specific assay depths. Different genes, sgRNAs, editors and delivery strategies may behave very differently. The authors emphasize the need for more loci, larger cohorts and broader safety assays.
Fifth, the study includes important genome-editing stakeholders, including disclosed commercial and patent interests around base editing and prime editing involving David R. Liu. This does not weaken the data, but it reinforces the need for independent replication when judging platform safety and generalizability.
The next questions are compensation, scope and safety
The most direct next question is why primitive endoderm can persist after NANOG loss in human embryos. Do FGF18, IGF1 or other RTK-signalling components truly compensate for FGF4? Is this only expression correlation, or is it functionally required? That will require more precise combinatorial perturbation.
A second direction is to expand the set of developmental regulators. NANOG is a high-value starting point, but human embryo functional genetics becomes more transformative if it can systematically test nodes such as OCT4, SOX2, GATA6, TFAP2C and CDX2, while separating conserved functions from species-specific rewiring across mouse, human embryos and stem-cell models.
A third direction is platform safety. ABE8e showed fewer obvious genotoxicity issues here than Cas9 nuclease, but this is a starting point rather than an endpoint. The field needs higher-resolution on-target structural-variant detection, genome-wide off-target profiling, RNA off-target assessment, long-term developmental-potential assays and direct comparisons among editor variants in human embryos.
A fourth direction is ethics and governance. Even if base editing makes human embryo functional studies more precise, clinical heritable genome editing cannot follow automatically from technical feasibility. The paper itself emphasizes that any clinical translation would require broad public debate, rigorous ethical oversight and societal support.
Yang’s signal rating: High
Axis 1, signal strength: High. The paper is strong on both the technical and biological axes: technically, ABE8e provides a cleaner route for human embryo gene perturbation than DSB-forming nucleases; biologically, the study directly shows that NANOG is required for human epiblast specification and reveals a human-mouse divergence in primitive endoderm fate.
Axis 2, maturity: Medium-Low. This remains basic research and platform validation. Sample scarcity, one-locus scope, mosaicism and safety generalizability are unresolved; the work is appropriate for advancing human developmental functional genetics, not for supporting clinical embryo editing.
One-sentence summary: The value of this paper is that base editing moves human embryo biology from seeing where genes are expressed to directly testing what they do, while reminding us that mouse embryos cannot substitute for human embryos themselves.